|
|
Spliceosoma |
|||||
|
Molecola del mese di maggio 2020 La microscopia elettronica criogenica ci ha rivelato come gli spiceosomi tagliano e ricuciono l'RNA messaggero.  Introduzione IntroduzioneNiente è mai semplice in biologia. I meccanismi fondamentali della trasmissione dell'informazione genetica sono stati scoperti studiando i batteri e mostravano che il DNA è trascritto in RNA messaggero, il quale poi è tradotto in proteine. Quando i ricercatori hanno cominciato a studiare organismi più evoluti, però, le cose si sono rivelate molto più complesse. I geni che si trovano nel nostro DNA sono in generale da quattro a dieci volte più lunghi delle catene di mRNA che vengono tradotte in proteine. Studiando il genoma si è visto che i tratti dei geni che codificano per le proteine sono suddivisi in più sequenze, chiamate esoni, separate da lunghi tratti non codificanti, chiamati introni. Il numero di esoni in cui è suddivisa l'informazione di una proteina è molto variabile: per esempio, il gene della nostra emoglobina (mdm 5-2003) è separato in tre sequenze, mentre il gene della nostra proteina più grande, la titina, è diviso in 365 sequenze. Questa modularità ha un grande vantaggio: le cellule possono tagliare e ricucire le sequenze copiate da un gene in più modi e così, con lo stessa sequenza originale di DNA, si possono sintetizzare più proteine capaci di svolgere funzioni un po' diverse tra loro. La reazione di splicing Gli spliceosomi tagliano la lunga catena di pre-mRNA appena trascritta dal gene e la riconnettono per produrre una catena più corta di mRNA maturo che poi è usata nella sintesi di proteine. Gli spliceosomi sono complesse macchine molecolari formate da più di 100 catene proteiche e 5 piccoli segmenti di RNA nucleare. Il complesso viene costruito passo-passo aggiungendo o rimuovendo subunità per realizzare i vari passaggi della reazione di splicing. Nel primo passaggio vengono riconosciuti e avvicinati tra loro i siti di splicing della catena di pre-mRNA. Poi il sito di splicing 5' è tagliato e l'estremità dell'introne viene legata ad uno speciale nucleotide di adenina al centro dello stesso introne, creando un anello. Infine l'altro sito di splicing, 3', è tagliato, le estremità dei due esoni si legano tra loro e l'introne che contiene l'anello (lazo) è rilasciato. Strutture dinamiche La microscopia elettronica criogenica è stata usata per esplorare i vari passaggi della reazione promossa dallo spliceosoma. Questo è un enzima estremamente dinamico quindi le strutture ottenute contengono solo le parti più rigide del complesso e dobbiamo usare un po' di immaginazione per riempire i vuoti lasciati dalle porzioni troppo flessibili per essere registrate. La struttura qui sopra (file PDB 3jb9) mostra lo spliceosoma di lievito alla fine della reazione di splicing, quando i due esoni connessi tra loro sono già stati rilasciati e l'introne con l'anello a forma di lazo è ancora legato. Con la microscopia elettronica criogenica sono state ottenute strutture che mostrano le varie fasi dello splicing dell'mRNA comprese quelle che coinvolgono gli spliceosomi più complessi umani e così si sono chiariti i dettagli strutturali dell'intero processo. 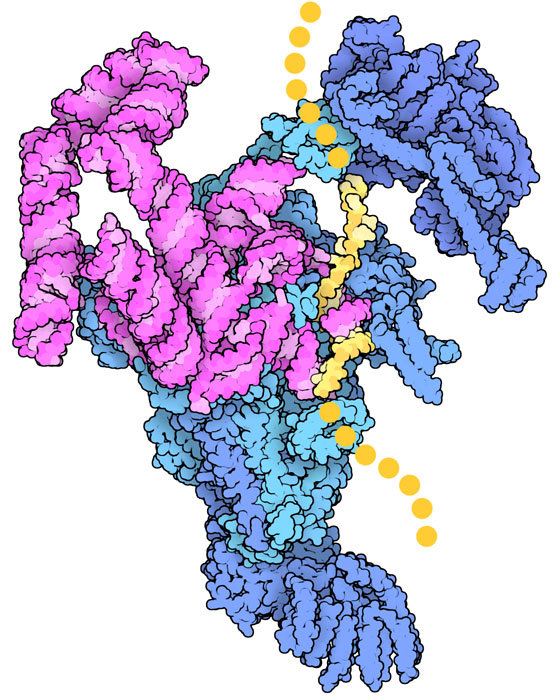 Il
ruolo dell'RNA Il
ruolo dell'RNALa maggior parte del lavoro nello spliceosoma è realizzato da piccole catene di RNA nucleare. Qui a lato si vede il complesso E dello spliceosoma di lievito (file PDB 6n7p) che mostra l'inizio del processo quando viene riconosciuto il primo dei due siti di splicing. L'RNA dello spliceosoma (magenta) si sta appaiando con il pre-mRNA (giallo) e, con l'aiuto delle proteine dell'enzima (blu), sta leggendo la sequenza del sito di splicing. A questo nucleo centrale, si uniscono poi altre proteine e altre catene di RNA per costruire la struttura completa dello spliceosoma. Queste catene di RNA eseguono le reazioni di taglio e ricucitura del pre-mRNA con l'aiuto di due ioni magnesio. Sorprendentemente esistono alcuni autosplicing mRNA (mdm 5-2005) che sono in grado di tagliare e ricucire la propria stessa catena sfruttando meccanismi simili, senza l'aiuto di spliceosomi. Collegamento col cancro  Il
funzionamento difettoso degli spliceosomi può dare origine a più
di 200 malattie, per questo è studiato così intensamente.
Per esempio, uno splicing errato produce una forma difettosa della proteina
di segnalazione Rac1 che ha un ruolo importante nello sviluppo
di alcuni tipi di cancro. Similmente alla proteina Ras
(mdm 04-2012), anche Rac1 è un interruttore molecolare che usa
GTP e GDP per accendere e spegnere i segnali di crescita e proliferazione
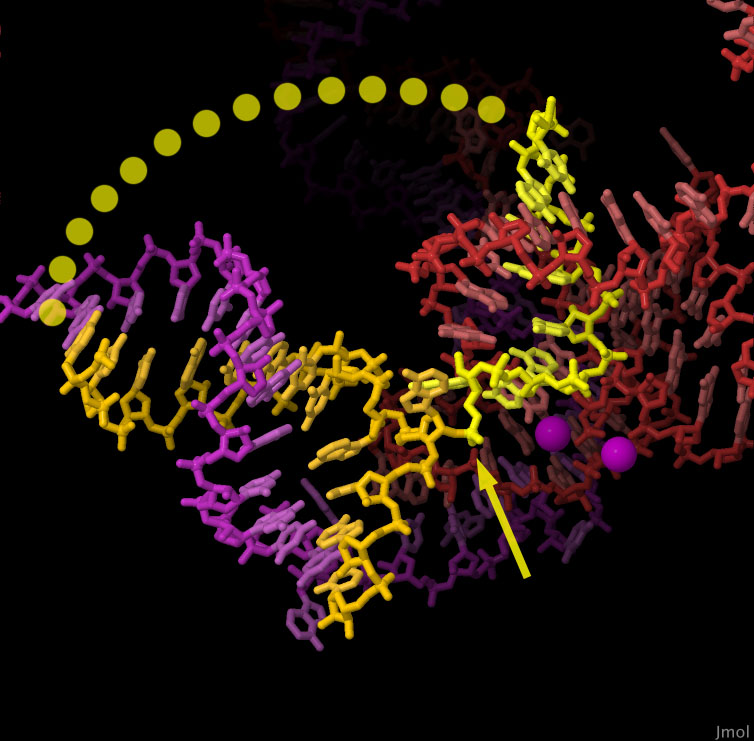
cellulare. Normalmente Rac1 (mostrata qui a fianco, file PDB 1mh1)
è codificata da 6 esoni, le corrispondenti sequenze di amminoacidi
nella proteina sono mostrate di colori diversi. La forma errata Rac1b
(file PDB 1ryf) è prodotta da un
mRNA con un esone in più nel centro della catena, la proteina così
ha un segmento extra (verde) che in parte sporge dalla struttura con un
tratto flessibile mostrato qui con dei pallini. Questo tratto modifica
in parte il comportamento di Rac1 che resta più a lungo nella fase
accesa. Alcune cellule cancerose producono molta Rac1b, che le aiuta a
crescere scavalcando i normali controlli. Il
funzionamento difettoso degli spliceosomi può dare origine a più
di 200 malattie, per questo è studiato così intensamente.
Per esempio, uno splicing errato produce una forma difettosa della proteina
di segnalazione Rac1 che ha un ruolo importante nello sviluppo
di alcuni tipi di cancro. Similmente alla proteina Ras
(mdm 04-2012), anche Rac1 è un interruttore molecolare che usa
GTP e GDP per accendere e spegnere i segnali di crescita e proliferazione
cellulare. Normalmente Rac1 (mostrata qui a fianco, file PDB 1mh1)
è codificata da 6 esoni, le corrispondenti sequenze di amminoacidi
nella proteina sono mostrate di colori diversi. La forma errata Rac1b
(file PDB 1ryf) è prodotta da un
mRNA con un esone in più nel centro della catena, la proteina così
ha un segmento extra (verde) che in parte sporge dalla struttura con un
tratto flessibile mostrato qui con dei pallini. Questo tratto modifica
in parte il comportamento di Rac1 che resta più a lungo nella fase
accesa. Alcune cellule cancerose producono molta Rac1b, che le aiuta a
crescere scavalcando i normali controlli.
Esplorando la struttura
|
||||||
|
|
||||||